

School of Psychology, University of Liverpool, Liverpool, UK
t.c.kirkham@liverpool.ac.uk

Introduction
Key Points
- CB1 agonists stimulate eating.
- CB1 antagonists suppress food intake.
- Endocannabinoids mediate specific motivational aspects of appetite.
- Endocannabinoid activity is linked to:
- Increased salience and incentive value of food and food-related stimuli through activation of mesolimbic dopamine incentive ('wanting') circuits.
- Enhanced palatability/reward of food via modulation of nucleus accumbens shell circuitry and interactions with endogenous opioids.
- CB1 agonists promote hunger, food craving, anticipation of food pleasure, and heightened enjoyment of food.
- CB1 antagonists can reduce food craving and hunger.
- The endocannabinoid system is key to overconsumption and weight gain as a component of the biological systems that have evolved to ensure positive energy balance.
- Pharmacological modulation of brain endocannabinoid activity may permit effective modification of 'greedy' behaviours and increased restraint overeating in the obese.
The predominant models that guide current development of drugs designed to address obesity and its associated diseases concentrate on homeostatic regulation of energy balance and body weight. However, it is arguable that neither body weight nor food intake are regulated variables in the generally accepted physiological sense. The increasing prevalence of obesity-arising from the overconsumption of palatable, energy-dense foods and from sedentary lifestyles-indicates the failure of any efficient mechanisms to curtail energy intake with increasing adiposity and of any effective break on the accumulation of body weight once the much vaunted 'set points' for weight or adiposity are attained. Rather, the mechanisms that underlie hunger, and particularly food-craving and hedonic responses to food, are considerably greater influences on the frequency, quantity, and variety of consumption. Consequently, should we wish to develop pharmaceutical interventions to restrict food intake, these positive motivational factors represent crucial targets for investigation and are likely to provide more effective therapies than agents that aim to reinforce putative inhibitory 'satiety signals'. The endocannabinoids appear to be critical to the normal biopsychological mechanisms that create appetite and stimulate eating, specifically contributing to incentive processes and the hedonic evaluation of food stimuli.
State of the Art
 [Click to enlarge]
[Click to enlarge]
The well-documented appetite-stimulating actions of Cannabis sativa result from agonist actions of phytocannabinoids, such as delta-9-tetrahydrocannabinol (THC), at CB1 receptors. That these actions reflect a physiological role of endocannabinoids in appetite control was confirmed by the demonstration in animal models that 1-CB1 blockade suppresses food intake and 2-the endogenous CB1 ligands anandamide, 2-arachidonoylglycerol (2-AG), and noladin ether all promote eating [1-4]. More detailed behavioural analysis indicates that endocannabinoids specifically modulate food wanting and liking. Thus, CB1 agonists and antagonists respectively increase or reduce the amount of effort an animal will expend to obtain food [5-7]. Additionally, CB1 knockout mice exhibit lower levels of responding for sweet food than wild-type mice [8]. THC, 2-AG, or anandamide advance the onset of meals, inducing eating even in fully-satiated animals [9-11]. The orexigenic actions of cannabinoids resemble the changes that occur with food deprivation, and regional brain levels of anandamide and 2-AG increase after fasting [12].
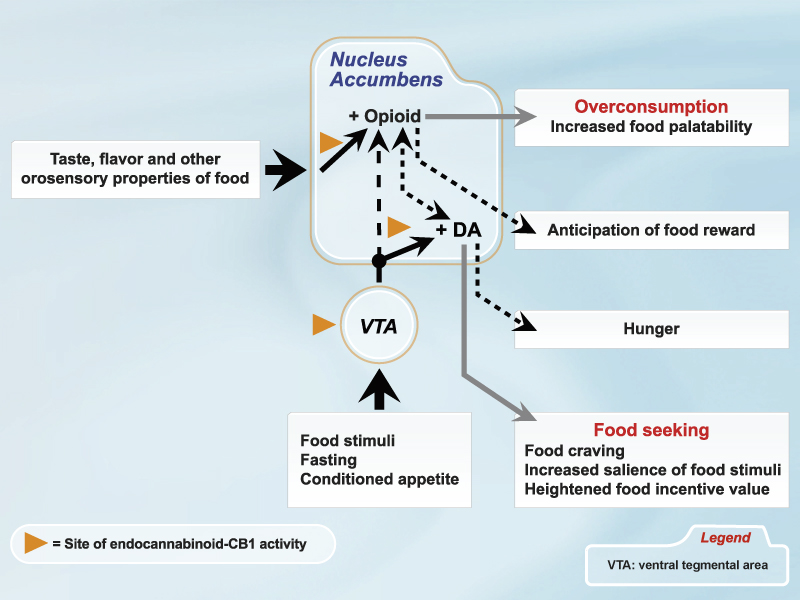
In humans, a principal effect of THC is the amplification of preprandial hunger [13]. Conversely, rimonabant selectively lowers hunger and desire to eat at the start of a meal, while having no effect on postmeal ratings of hunger or fullness. Importantly, with repeated administration, rimonabant reduces the frequency and strength of food cravings [14]. These data are compatible with the known effects of CB1 agonists and antagonists on mesolimbic dopaminergic neurons that subserve incentive motivation (Figure). For example, the accumbens dopamine release that is provoked by presentation of a novel, palatable food is blocked by rimonabant [15]. Overall, the data imply that endocannabinoids may be essential to the orientation to motivationally significant stimuli, the attribution of incentive salience and reward anticipation, and the elicitation of food seeking and eating initiation.
Endocannabinoids also appear to have a secondary role in mediating the liking of food. Positive hedonic reactions to sweet fluids are respectively enhanced or diminished by CB1 agonists and antagonists [16-18]. Moreover, the nucleus accumbens shell that mediates palatability responses is highly sensitive to the stimulatory actions of endocannabinoids. Anandamide and 2-AG are effective orexigens in this region, as are agents that increase endocannabinoid levels by blocking their enzymatic breakdown or reuptake [19]. Intra-accumbens administration of anandamide specifically enhances the hedonic impact of sweet taste [20], while accumbens CB1 receptors are down-regulated in rats that overconsume palatable food supplements [21], which is consistent with increased endocannabinoid activity.
Opioid receptor agonists and antagonists respectively increase or reduce food intake by altering the hedonic evaluation of foods [22-26]. There is now convincing evidence for interactions between endocannabinoids and endogenous opioids in relation to feeding. Thus, THC hyperphagia is attenuated by sub-anorectic doses of naloxone [27], and THC stimulates beta-endorphin release in the accumbens [28]. Importantly, the facilitatory effects of both CB1 and opioid receptor agonists on responding for palatable ingesta are reversed by either rimonabant and naloxone [5, 6]. Moreover, low doses of rimonabant and opioid antagonists that are behaviourally inactive when administered singly, combine synergistically to produce a profound anorectic action when co-administered [29, 30]. As with anandamide, administration of morphine into the accumbens shell increases the liking of sweet solutions, with a very close correspondence between opioid- and cannabinoid-sensitive sites [20, 31].
Independent manipulations of endocannabinoid or opioid processes produce distinct behavioural/motivational consequences, indicating that cannabinoids primarily affect appetitive processes while opioids mainly influence consummatory processes. We suggest that endocannabinoids principally mediate the motivational processes that drive us to eat, but-through interactions with opioid peptide systems-may also contribute significantly to the hedonic evaluation of foods during eating. Arguably, endocannabinoid-opioid activity underlies food craving, the anticipation of delight from eating, and the actual experience of pleasure derived from the sensory properties of food [9].
Priorities for Future Studies
As this paper indicates, central endocannabinoid systems are implicated in the principal psychological processes that govern eating motivation and may represent critical components of the mechanisms that lead us to overconsume, a major contributor to weight gain. As such, the endocannabinoids are potentially important therapeutic targets for pharmacological treatments designed to modify eating behaviours and attitudes/responsiveness to foods. Modification of endocannabinoid activity or blockade of CB1 receptors may allow us to limit our susceptibility to the temptations of food and to learn to restrain our excessive appetites. As these factors contribute more than any others to the development of obesity, there is an urgent need to define the psychological consequences of CB1 receptor manipulations in human studies. Insights obtained from the exploration of the subjective effects of CB1 ligands would shed important light on the true physiological role of endocannabinoids in appetite control.
References
- Arnone M, Maruani J, Chaperon F, et al. Selective inhibition of sucrose and ethanol intake by SR 141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology (Berl) 1997; 132: 104-6.
- Jamshidi N and Taylor DA. Anandamide administration into the ventromedial hypothalamus stimulates appetite in rats. Br J Pharmacol 2001; 134: 1151-4.
- Kirkham TC, Williams CM, Fezza F, et al. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: stimulation of eating by 2-arachidonoyl glycerol. Br J Pharmacol 2002; 136: 550-7.
- Williams CM and Kirkham TC. Anandamide induces overeating: mediation by central cannabinoid (CB1) receptors. Psychopharmacology (Berl) 1999; 143: 315-7.
- Avraham Y, Menachem AB, Okun A, et al. Effects of the endocannabinoid noladin ether on body weight, food consumption, locomotor activity, and cognitive index in mice. Brain Res Bull 2005; 65: 117-23.
- Gallate JE and McGregor IS. The motivation for beer in rats: effects of ritanserin, naloxone and SR 141716. Psychopharmacology (Berl) 1999; 142: 302-8.
- Gallate JE, Saharov T, Mallet PE, et al. Increased motivation for beer in rats following administration of a cannabinoid CB1 receptor agonist. Eur J Pharmacol 1999; 370: 233-40.
- Solinas M and Goldberg SR. Motivational effects of cannabinoids and opioids on food reinforcement depend on simultaneous activation of cannabinoid and opioid systems. Neuropsychopharmacology 2005; 30: 2035-45.
- Kirkham TC and Williams CM. Endocannabinoids: neuromodulators of food craving? In: Hetherington M (ed.), Food Cravings and Addiction, Leatherhead Publishing, Surrey, pp. 85-120, 2001.
- Sanchis-Segura C, Cline BH, Marsicano G, et al. Reduced sensitivity to reward in CB1 knockout mice. Psychopharmacology (Berl) 2004; 176: 223-32.
- Williams CM and Kirkham TC. Observational analysis of feeding induced by Delta9-THC and anandamide. Physiol Behav 2002; 76: 241-50.
- Kirkham TC. Endocannabinoids in the regulation of appetite and body weight. Behav Pharmacol 2005; 16: 297-313.
- Townson AL. Acute effects of oromucosal administration of a phytocannabinoid, D9-THC, on appetite and mood in healthy volunteers. Unpublished PhD Thesis, University of Liverpool, 2007.
- Blundell JE, Jebb S, Stubbs RJ, et al. Effect of rimonabant on energy intake, motivation to eat and body weight with or without hypocaloric diet: the REBA study. Obes Rev 2006; 7 (Suppl 2): 104.
- Melis T, Succu S, Sanna F, et al. The cannabinoid antagonist SR 141716A (Rimonabant) reduces the increase of extra-cellular dopamine release in the rat nucleus accumbens induced by a novel high palatable food. Neurosci Lett 2007; 419: 231-5.
- Higgs S, Williams CM and Kirkham TC. Cannabinoid influences on palatability: microstructural analysis of sucrose drinking after delta(9)-tetrahydrocannabinol, anandamide, 2-arachidonoyl glycerol and SR141716. Psychopharmacology (Berl) 2003; 165: 370-7.
- Jarrett MM, Limebeer CL and Parker LA. Effect of Delta9-tetrahydrocannabinol on sucrose palatability as measured by the taste reactivity test. Physiol Behav 2005; 86: 475-9.
- Jarrett MM, Scantlebury J and Parker LA. Effect of delta9-tetrahydrocannabinol on quinine palatability and AM251 on sucrose and quinine palatability using the taste reactivity test. Physiol Behav 2007; 90: 425-30.
- Soria-Gomez E, Matias I, Rueda-Orozco PE, et al. Pharmacological enhancement of the endocannabinoid system in the nucleus accumbens shell stimulates food intake and increases c-Fos expression in the hypothalamus. Br J Pharmacol 2007; 151: 1109-16.
- Mahler SV, Smith KS and Berridge KC. Endocannabinoid hedonic hotspot for sensory pleasure: anandamide in nucleus accumbens shell enhances 'liking' of a sweet reward. Neuropsychopharmacology 2007; 32: 2267-78.
- Harrold JA, Elliott JC, King PJ, et al. Down-regulation of cannabinoid-1 (CB-1) receptors in specific extrahypothalamic regions of rats with dietary obesity: a role for endogenous cannabinoids in driving appetite for palatable food? Brain Res 2002; 952: 232-8.
- Bodnar RJ. Endogenous opioids and feeding behavior: a 30-year historical perspective. Peptides 2004; 25: 697-725.
- Cooper SJ and Kirkham TC. Opioid mechanisms in the control of food consumption and taste preferences. In: Herz A, Akil A, Simon EJ (eds.), Handbook of Experimental Pharmacology, Volume 104 (Part II), Springer-Verlag, Berlin, pp. 239-263, 1993.
- Drewnowski A, Krahn DD, Demitrack MA, et al. Taste responses and preferences for sweet high-fat foods: evidence for opioid involvement. Physiol Behav 1992; 51: 371-9.
- Kirkham TC and Cooper SJ. Naloxone attenuation of sham feeding is modified by manipulation of sucrose concentration. Physiol Behav 1988; 44: 491-4.
- Yeomans MR and Gray RW. Selective effects of naltrexone on food pleasantness and intake. Physiol Behav 1996; 60: 439-46.
- Williams CM and Kirkham TC. Reversal of delta 9-THC hyperphagia by SR141716 and naloxone but not dexfen-fluramine. Pharmacol Biochem Behav 2002; 71: 333-40.
- Solinas M, Zangen A, Thiriet N, et al. Beta-endorphin elevations in the ventral tegmental area regulate the discriminative effects of Delta-9-tetrahydrocannabinol. Eur J Neurosci 2004; 19: 3183-92.
- Chen RZ, Huang RR, Shen CP, et al. Synergistic effects of cannabinoid inverse agonist AM251 and opioid antagonist nalmefene on food intake in mice. Brain Res 2004; 999: 227-30.
- Kirkham TC and Williams CM. Synergistic efects of opioid and cannabinoid antagonists on food intake. Psychopharmacology (Berl) 2001; 153: 267-70.
- Pecina S and Berridge KC. Opioid site in nucleus accumbens shell mediates eating and hedonic 'liking' for food: map based on microinjection Fos plumes. Brain Res 2000; 863: 71-86.
