

Endocrinology Unit and Center of Applied Biomedical Research (C.R.B.A.), Department of Internal Medicine and Gastroenterology, S. Orsola-Malpighi Hospital, Alma Mater Studiorum University of Bologna, Bologna, Italy
uberto.pagotto@unibo.it

Introduction
Key Points
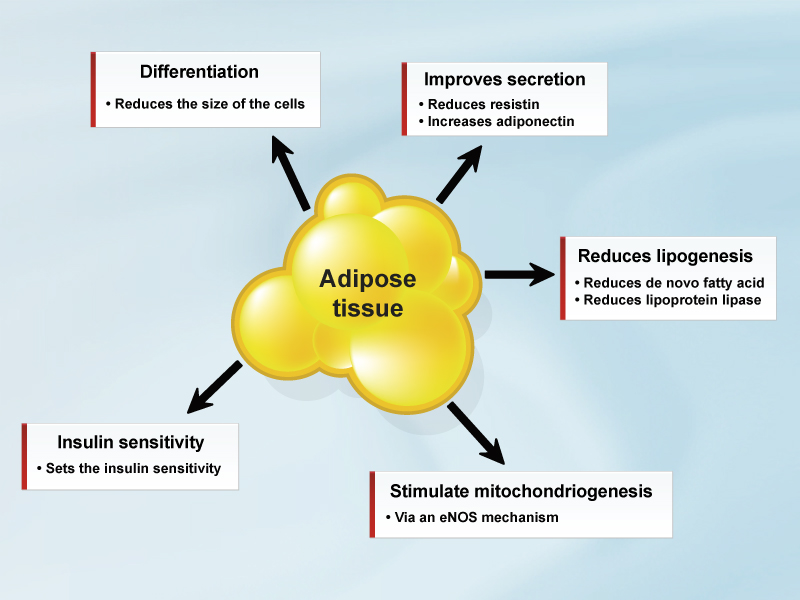
- Endocannabinoids have emerged as an important modulator of several functions of adipose tissue, including cell proliferation, differentiation, secretion, lipogenesis, mitochondriogenesis, and insulin sensitivity.
- Overactivation of the ECS is associated with the development and/or maintenance of obesity.
- A novel class of drugs, the CB1 antagonists, have been shown to have a beneficial metabolic effect beyond weight loss in obese patients. This effect can be directly attributed to the blockade of the ECS at the level of adipose tissue.
Historically, the endocannabinoid system has been thought to affect energy metabolism by its motivational and orexigenic neuronal effect. However, the weight loss induced by the antagonization of CB1 receptor has only recently been linked to a reduction in caloric intake, which remains a mechanism limited in time. In addition, this weight loss has been shown to be mainly controlled by sustained food intake-independent mechanisms that are more related to energy dispersion. The finding of a weight loss beyond the reduction in food intake led to the examination of the putative role of the endocannabinoid system (ECS) in the various organs implicated in energy dispersion. Very soon, it appeared evident that adipose tissue represents one of the most important targets to examine. This paper will describe the recent advancements in the interaction between adipose tissue and the ECS.
State of the Art
 [Click to enlarge]
[Click to enlarge]
The relevant role of endocannabinoids in adipocyte physiology is highlighted by the evidence of the presence of a functional ECS in the adipocytes. In fact, these cells not only express the CB1 receptor but are also endowed with the full biochemical machinery to synthesize and degrade endocannabinoids [1-9]. Although recent documentation of CB2 expression in adipocytes is noteworthy, its functional role is not well understood at present [6, 7, 9].
The expression of the CB1 receptor is more prominent in mature adipocytes than in pre-adipocytes [3, 6, 7]. Through these dynamic changes in CB1 profile expression, endocannabinoids promote adipocyte growth and differentiation, a function resulting from a cross-talk with the peroxisome proliferator-activated receptor-γ (PPAR-γ) [10]. Even more importantly, the CB1 receptor has been shown to be differently expressed in different compartments of adipose tissue, such as adipocytes derived from the omental and the subcutaneous fat pad, respectively. This finding makes it possible to speculate that the ECS, similar to other hormones and peptides, may act differently in these two districts [7].
The role of the endocannabinoids is not limited to increasing mature adipocyte proliferation. As in the brain, CB1 activation in adipose tissue acts to increase energy storage. Endocannabinoids influence a number of intracellular mechanisms to stimulate lipogenesis by inducing triglyceride accumulation through the inhibition of adenylate cyclase [4, 11] and the consequent reduction of lipolysis and via activation of lipoprotein lipase to provide exogenous fatty acids for the adipocytes [2]. In addition, endocannabinoids stimulate de novo lipogenesis in adipocytes by increasing the expression and activity of enzymes involved in fatty acid and triglyceride biosynthesis [12]. Finally, CB1 activation in adipocytes modulates insulin signalling and glucose uptake in order to increase energy storage [7, 13]. Recent data seems to show that endocannabinoids determine the sensitivity of the insulin response in adipocytes through an involvement of Akt [13].
In the past few years, it has been established that adipose tissue may also act as an endocrine organ. In fact, adipocytes express and secrete a number of adipokines that may deeply influence local adipocyte biology as well as systemic metabolism at sites as diverse as the brain, liver, muscle, pancreatic β-cells, gonads, lymphoid organs, and systemic vasculature [14]. Endocannabinoids have been recognized as important players in the regulation of adipokine secretion in down-regulating adiponectin [1, 3, 6, 11] and stimulating visfatin secretion [11, 15], contributing to the impairment of insulin sensitivity and a decrease in glucose uptake in skeletal muscle.
Recently, it has been found that the administration of CB1 antagonists first in diet-induced obese rodents and after in obese subjects induced an increase in energy expenditure [16-18]. These findings, together with an analysis of the intra-adipocyte gene modulations induced by CB1 blockade chronic treatment, indicated that the drug-induced reduction of adipose mass is attributable to increased energy expenditure, mainly through futile cycling (calcium and substrate) [11]. Rimonabant treatment also altered gene expression, which favoured energy dissipation through mitochondrial heat production in brown adipose tissue [11]. Several reports have documented a direct role of the ECS in the modulation of proteins involved in thermogenesis [1, 11]. Treatment of differentiated brown adipocytes with a CB1 agonist decreased the expression of uncoupling protein 1 [11, 15]. However, unlike with rodents, the role of brown adipose tissue in humans is not yet well established. Physiological and pharmacological stimuli may be capable of trans-differentiating white adipocytes into brown adipocytes in humans. Therefore, CB1 receptor antagonist treatment may lead to an eventual increase in energy expenditure by increasing brown adipocytes. Genetic and pharmacological blockade of the cannabinoid CB1 receptor increases mitochondrial biogenesis in white and brown adipocytes [19]. Treatment of adipocytes with rimonabant, a selective antagonist of CB1 receptors, increases mitochondrial biogenesis genes, including PGC-1a and Tfam. This possibly implies that SR141716 is antagonizing an endocannabinoid tone present in cultured fat cells. Moreover, the genetic CB1 receptor blockade in vivo increases eNOS expression and mitochondrial biogenesis both in whole white adipose tissue and isolated mature white adipocytes, and this is accompanied by prevention of high fat diet-induced fat accumulation. This might increase oxidative metabolism in white adipocytes by counteracting the inhibitory effects of endocannabinoids, whose levels are increased in fat tissues of obese rodents and humans [19].
A possible mediator of these functions may be the AMPK system, which acts as a fuel sensor to regulate energy balance both at the cellular level and within the whole body, by inhibiting anabolic pathways and stimulating catabolic processes in order to increase the ATP/AMP ratio. Endocannabinoids act to decrease AMPK activity in the adipose tissue, contributing to an increase in adiposity and lipogenesis and resulting in decrease in energy expenditure [19]. Accordingly, induction of AMPK activity has been found to increase mitochondrial content in adipocytes. Thus, the enhanced ß-oxidation of free fatty acids elicited by AMPK activation and by eNOS-dependent mitochondrial biogenesis might functionally link rimonabant treatment to its anti-obesity effects.
As mentioned before, a close association between the development of obesity and a simultaneous overactivation of the ECS in the adipose tissue, expressed as a rise in endocannabinoid production or an increase in CB1 receptor expression, has been shown [6, 7]. Significantly higher levels of 2-arachidonoylglycerol (2-AG), but not anandamide, have been detected in the visceral (intra-abdominal), but not subcutaneous, fat of obese patients [7]. A similar pathological increase in endo-cannabinoids has been detected in mice. However, in rodents, anandamide and not 2-AG seem to be elevated [12]. Importantly, this increased intra-adipocitic tone of endocannabinoids has been demonstrated to be under the control of hypothalamic leptin signalling [12].
Increased levels of plasma 2-AG were present in patients with visceral obesity compared to patients with subcutaneous obesity and compared to lean controls [20-22]. The increase in 2-AG was also shown to positively correlate with some important cardiometabolic risk factors, such as body mass index, waist circumference, fasting plasma triglyceride and insulin levels, low HDL cholesterol, and adiponectin levels [21].
Altogether, this data enables us to include the CB1 receptor and the endocannabinoids in the group of agents that play a determinant role in the physiology and the pathophysiology of these cells (figure).
References
- Bellocchio L, Cervino C, Vicennati V, et al. Cannabinoid type 1 receptor: another arrow in the adipocytes' bow. J Neuroendocrinol 2008; 20 Suppl 1: 130-8.
- Cota D, Marsicano G, Tschop M, et al. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J Clin Invest 2003; 112: 423-31.
- Gary-Bobo M, Elachouri G, Scatton B, et al. The cannabinoid CB1 receptor antagonist rimonabant (SR141716) inhibits cell proliferation and increases markers of adipocyte maturation in cultured mouse 3T3 F442A preadipocytes. Mol Pharmacol 2006; 69: 471-8.
- Gasperi V, Fezza F, Pasquariello N, et al. Endocannabinoids in adipocytes during differentiation and their role in glucose uptake. Cell Mol Life Sci 2007; 64: 219-29.
- Gonthier MP, Hoareau L, Festy F, et al. Identification of endocannabinoids and related compounds in human fat cells. Obesity (Silver Spring) 2007; 15: 837-45.
- Matias I, Gonthier MP, Orlando P, et al. Regulation, function, and dysregulation of endocannabinoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia. J Clin Endocrinol Metab 2006; 91: 3171-80.
- Pagano C, Pilon C, Calcagno A, et al. The endogenous cannabinoid system stimulates glucose uptake in human fat cells via phosphatidylinositol 3-kinase and calcium-dependent mechanisms. J Clin Endocrinol Metab 2007; 92: 4810-9.
- Spoto B, Fezza F, Parlongo G, et al. Human adipose tissue binds and metabolizes the endocannabinoids anandamide and 2-arachidonoylglycerol. Biochimie 2006; 88: 1889-97.
- Roche R, Hoareau L, Bes-Houtmann S, et al. Presence of the cannabinoid receptors, CB1 and CB2, in human omental and subcutaneous adipocytes. Histochem Cell Biol 2006; 126: 177-87.
- Bouaboula M, Hilairet S, Marchand J, et al. Anandamide induced PPARgamma transcriptional activation and 3T3-L1 preadipocyte differentiation. Eur J Pharmacol 2005; 517: 174-81.
- Jbilo O, Ravinet-Trillou C, Arnone M, et al. The CB1 receptor antagonist rimonabant reverses the diet-induced obesity phenotype through the regulation of lipolysis and energy balance. Faseb J 2005; 19: 1567-9.
- Buettner C, Muse ED, Cheng A, et al. Leptin controls adipose tissue lipogenesis via central, STAT3-independent mechanisms. Nat Med 2008; 14: 667-75.
- Motaghedi R and McGraw TE. The CB1 endocannabinoid system modulates adipocyte insulin sensitivity. Obesity (Silver Spring) 2008; 16: 1727-34.
- Ahima RS. Adipose tissue as an endocrine organ. Obesity (Silver Spring) 2006; 14 Suppl 5: 242S-9S.
- Perwitz N, Fasshauer M and Klein J. Cannabinoid receptor signaling directly inhibits thermogenesis and alters expression of adiponectin and visfatin. Horm Metab Res 2006; 38: 356-8.
- Addy C, Wright H, Van Laere K, et al. The acyclic CB1R inverse agonist taranabant mediates weight loss by increasing energy expenditure and decreasing caloric intake. Cell Metab 2008; 7: 68-78.
- Dernbach H, Haller J, Kaulisch T, et al. Effects of the CB1 receptor antagonist SR141716A (rimonabant) on energy expenditure and respiratory quotient in diet-induced obese rats. Obes Rev 2005; 6: 172 (abstract).
- Herling AW, Kilp S, Elvert R, et al. Increased energy expenditure contributes more to the body weight-reducing effect of rimonabant than reduced food intake in candy-fed wistar rats. Endocrinology 2008; 149: 2557-66.
- Tedesco L, Valerio A, Cervino C, et al. Cannabinoid type 1 receptor blockade promotes mitochondrial biogenesis through endothelial nitric oxide synthase expression in white adipocytes. Diabetes 2008; 57: 2028-36.
- Bluher M, Engeli S, Kloting N, et al. Dysregulation of the peripheral and adipose tissue endocannabinoid system in human abdominal obesity. Diabetes 2006; 55: 3053-60.
- Coté M, Matias I, Lemieux I, et al. Circulating endocannabinoid levels, abdominal adiposity and related cardiometabolic risk factors in obese men. Int J Obes (Lond) 2007; 31: 692-9.
- Engeli S, Bohnke J, Feldpausch M, et al. Activation of the peripheral endocannabinoid system in human obesity. Diabetes 2005; 54: 2838-43.
